
Расшифровывая язык пчел (Нобелевская лекция К. фон Фриша)
Нобелевская лекция 12 декабря 1973 года
Перевод с английского: Максим Мороз.
КАРЛ ФОН ФРИШ
Университет Мюнхена, Федеративная Республика Германия
Еще 60 лет назад многие биологи считали, что пчелы и другие насекомые совершенно не различают цвета. Я не мог в это поверить. Яркость цветов может объясняться только адаптацией к посетителям, различающим цвет. Это положило начало экспериментам с цветовым восприятием у пчел (I). На столе во дворе я поместил лист цветной бумаги между листами бумаги, окрашенными в различные оттенки серого цвета, и на цветной лист — маленькую стеклянную тарелку с сахарным сиропом*. Пчелы с близлежащего улья могли научиться распознавать этот цвет и демонстрировали способность отличать его от оттенков серого. Во избежание сбора пчел в чрезмерных количествах, я установил перерывы между кормлениями. После этих перерывов, только единичные пчелы-разведчики прилетали к пустой тарелке и возвращались домой; стол для кормления оставался пустым. Однако, если пчела-разведчик обнаруживала наполненную миску и успешно возвращалась домой, уже через несколько минут возвращалась вся группа фуражиров (пчел-сборщиц). Она сообщала о своей находке рою? Этот вопрос стал отправной точкой для дальнейших исследований.
Так как нет возможности наблюдать поведение пчел-сборщиц после их возвращения в улей, небольшая колония была помещена для наблюдений в улей со стеклянными окнами, а рядом была установлена тарелка для кормления. Отдельные пчелы-сборщицы были отмечены цветными точками, пронумерованными по определенной системе. Теперь в наблюдательном улье можно было видеть поразительную картину: еще до того как возвращающиеся пчелы выдавали содержимое своего медового зобика другим пчелам, они бегали по сотам кругами малого радиуса, поочередно вправо и влево. Такой круговой танец заставлял пронумерованных пчел двигаться за ними для проведения новой экскурсии в место кормления.
Однако пчелы-фуражиры из одного улья не всегда летают к одному источнику кормления. Наблюдается формирование сборщиц в группы: одни могут собирать пыльцу с одуванчиков, другие с клевера, а третьи – с незабудок. Даже при наличии цветущих растений часто возможна нехватка пищи, что приводит к «перерыву в питании». Были ли пчелы, участвовавшие в эксперименте, способны оповещать тех же пчел-сборщиц, что были с ними у миски для кормления? Способны ли они распознавать друг друга индивидуально?
Чтобы ответить на этот вопрос, я установил два места для кормления, на которых по отдельности собирались две группы пчел из того же наблюдательного улья. Во время перерыва в питании обе группы оставались на сотах и смешивались между собой. После этого наполнялась одна из мисок для кормления. Пчелы, прилетающие от наполненной миски сообщали о ней своими танцами не только пчелам своей группы, но также и пчелам второй группы, которые реагировали вылетом на свое привычное место кормления, где они обнаруживали пустую миску.
Однако в естественной среде пчелы ищут не стеклянные миски, а цветы. Поэтому в эксперимент были внесены изменения; одна из двух групп пчел собирала пищу с цветков липы, а другая – с белой акации. Теперь картина изменилась: после перерыва в кормлении, при возвращении пчел с цветков липы снова вылетали только пчелы-сборщицы липы; при этом сборщицы белой акации не обращали на танцы сборщиц липы никакого внимания. С другой стороны, при успешном возвращении пчел с цветов акации, пчелы-сборщицы липы не выказывали никакого интереса к их танцам, при этом члены акациевой группы незамедлительно подбегали к ближайшей танцующей пчеле, следуя за ней, после чего вылетали из улья. Некоторые смышленые пчелы также научились использовать от случая к случаю оба источника пищи. Потом они могли высылать сборщиц акации после возвращения с цветков акации и сборщиц липы после посещения цветков липы. Однако пчелы не знали друг друга по отдельности. Оказалось, что решающее значение имеет аромат определенных цветов, исходящий от их тел. Для подтверждения этой теории использовались эфирные масла или синтетические ароматизаторы в местах кормления, которые дали тот же эффект.
При длительном кормлении, новые рекруты появлялись у источника пищи после старых сборщиц. Они также оповещались танцем. Однако как они находили свою цель?
В место кормления рядом с ульем было добавлено масло мяты перечной. Кроме того, миски с сахарным сиропом были установлены на небольшие листы картона в разных местах на лугу рядом; на некоторые листы наносилось масло мяты перечной, а на другие листы – другие эфирные масла. Результат был однозначным: через несколько минут после начала кормления рекруты из наблюдательного улья появились не только на месте кормления рядом с пчелами-сборщицами, но и на других чашах с мятой перечной, установленных на лугу, на некотором расстоянии. Однако другие чаши с нанесенным запахом оставались нетронутыми. Запах лаванды, фенхеля, масла тимьяна и т. п. не привлекал внимания пчел. Когда запах в месте кормления был заменен другим, соответственно изменилась цель роения рекрутов. Они руководствовались ароматом, который приносили на себе танцующие пчелы.
Запах — очень простой, но эффективный способ общения. Однако он приобретает полное значение только в сочетании с другим условием. В случае дефицита сахарного сиропа или его слабой концентрации, с определенного момента танцы замедляются и, наконец, прекращаются, при этом сбор может продолжаться. С другой стороны, чем слаще сахарный сироп, тем ярче и длиннее становятся различные танцы. Эффект рекламы таким образом усиливается, и его дополнительно увеличивает ароматическая железа в брюшке пчелы-сборщицы, которая активизируется после прибытия пчелы с богатого источника пищи. Она будто подает находящимся поблизости рекрутам сигнал «иди сюда!». У многих самок насекомых имеются ароматические железы, которые привлекают самцов. Однако рабочие пчелы являются просто «рабочими лошадками», у которых отсутствует функция размножения, и их ароматические железы служат для информирования своего сообщества.
Теперь давайте представим себе весенний луг. Самые разные растения цветут одновременно, производя нектар разной концентрации. Чем богаче и слаще его поток, тем более живым будет танец пчел, которые находят и посещают цветы определенного типа. Цветы с лучшим нектаром передают определенный аромат, который обеспечивает им наибольшую популярность у пчел. Таким простым способом природа регулирует движение в соответствии с законом спроса и предложения не только в пользу пчел, но и способствует опылению и урожайности семян растений, богатых нектаром. Это открывает новую и доселе неизвестную сторону биологической важности цветочного аромата. Его огромное разнообразие и строгая специфика видов формируют поистине чарующий язык ароматов.
Так всё было в 1923 году (2), и я полагал, что знаю язык пчел. После возобновления экспериментов 20 лет спустя я заметил, что самый красивый аспект от меня ускользнул. Тогда я впервые установил место для кормления на расстоянии нескольких сотен метров, а не рядом с ульем, и с удивлением обнаружил, что рекруты немедленно начали добывать пищу на таком большом расстоянии, не обращая внимания на чаши возле улья. Противоположное имело место, когда пчелы-сборщицы обнаруживали сахарный сироп, как и прежде, возле улья. Могли ли они получать сигналы о дистанции?
Из одного наблюдательного улья были сформированы две группы пчел-сборщиц. Одно место для кормления было расположено в 12 м от улья, другое — на расстоянии 300 м. Открыв наблюдательный улей, я был поражен, увидев, что все пчелы-сборщицы из места кормления поблизости совершали круговые танцы, в то время как пчелы с дальнего места для кормления совершали танцы с вилянием брюшком (рис.1). При пошаговом перемещении ближнего места для кормления на большее расстояние круговые танцы сменялись «виляющими» танцами при достижении расстояния около 50 м. Второе место кормления пошагово перемещали к улью, мимо первого места кормления, находящегося рядом с ульем. На том же критическом расстоянии около 50 м виляющие танцы сменились круговыми танцами (3,4). Мне давно были известны виляющие танцы у пчел, но я считал их типичными для сборщиц пыльцы. Причиной моей ошибки было то, что пчелы с пыльцевыми корзинами всегда прибывали с большего расстояния, чем мои сборщицы сахарного сиропа.
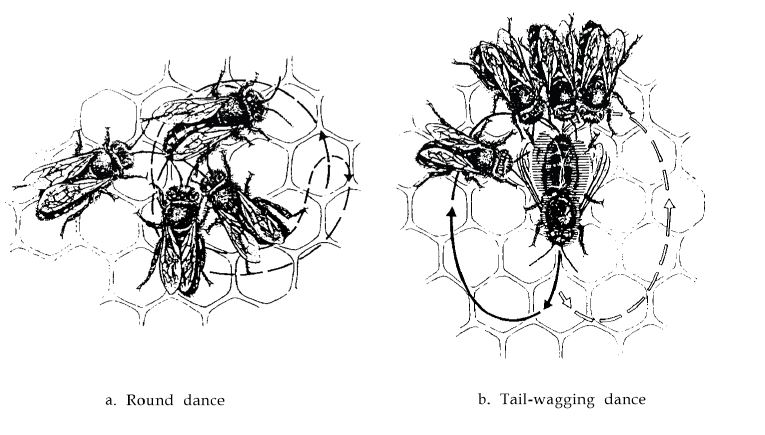
Рис. 1 Траектория движения пчелы во время (а) кругового танца и (б) танца с вилянием брюшком. Пчелы, которые следуют за танцующей пчелой, получают информацию.
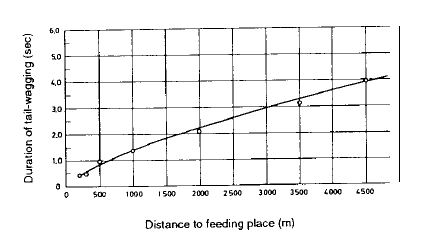
Таким образом, стало очевидным, и последующие эксперименты подтвердили (5), что круговой танец является сигналом, который символически приглашает членов улья к поиску в непосредственной близости от улья. Танец с вилянием брюшком призван отправить других пчел на большие расстояния, нередко на несколько километров. Сигнал «ближе 50 м» или «дальше 50 м» не сработал бы. Однако на самом деле темп виляющего танца изменяется пропорционально увеличению расстояния: его ритм уменьшается. Согласно современному уровню наших знаний, информация о дистанции полета соответствует длительности времени, необходимого для прохождения прямой части фигуры – танца в форме восьмерки в каждом повторе. Этот прямой отрезок четко обозначается танцевальными движениями брюшком и одновременно озвучивается (в прямом смысле) жужжанием (6, 7). Более длинные расстояния символически выражаются более продолжительным вилянием брюшком. Для расстояний от 200 до 4500 м продолжительность увеличивается от примерно 0,5 секунды до примерно 4 секунд (6, 8) (рис.2).
Distance to feeding place – Расстояние до места кормления (м)
Рис. 2. Продолжительность пробега с виляния брюшком для мест кормления, находящихся на разных расстояниях, на основе данных видеозаписей.
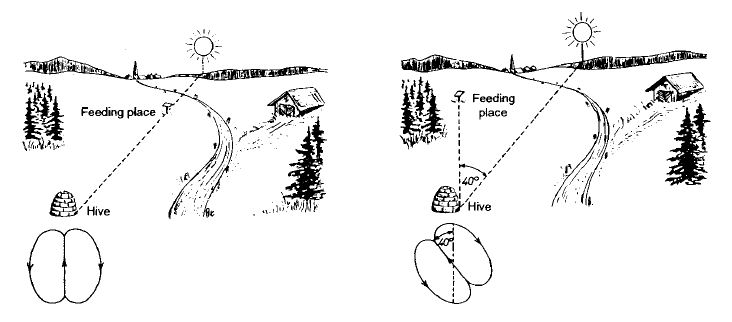
«Виляющий» танец указывает не только расстояние, но и направление к цели. В наблюдательном улье пчелы, прибывающие с одного места кормления, делают «виляющие» пробеги в одном и том же направлении, в то время как пробеги ориентированы иначе у пчел, прибывающих с других направлений. Однако направление пробегов с вилянием брюшком у пчел, прибывающих с одного места для кормления, не сохраняется постоянным. В течение дня направление меняется на тот же угол, что и угол, который в это время проходит солнце, но в противоположном направлении. Таким образом, танцующая пчела показывает другим пчелам направление к цели по отношению к положению солнца (5, 6). Те часы, которые я провел у наблюдательного улья, когда пчелы открыли мне этот секрет, останутся для меня незабываемыми. Завораживает то, что угол между положением солнца и траекторией танцующей пчелы к цели выражается пчелой в темноте улья на вертикальной поверхности соты как угловое отклонение от вертикали. Таким образом, пчела переносит угол в другую область чувственного восприятия. На рисунке 3 показан ключ к преобразованию. Если цель находится в направлении солнца, виляющие танец направлен вверх. Если цель находится в 40° влево от положения солнца, танцующая пчела смещает прямую траекторию на 40° влево от вертикали и т. д. (5,6). На соте члены улья двигаются за танцующей пчелой и поддерживают с ней тесный контакт, особенно на отрезках с вилянием брюшком, и получают предлагаемую информацию. Могут ли они следовать ей и с какой точностью?
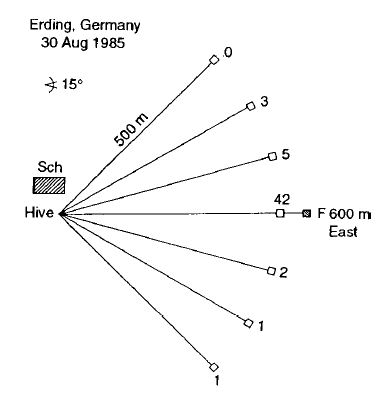
Для проверки указания направления мы воспользовались следующим методом (9). На определенном расстоянии от улья было установлено место для кормления, на котором на не ароматизированной платформе пронумерованные пчелы кормились сахарным раствором, который был настолько разбавлен, что они не исполняли танца в улье и потому не оповещали новичков. Только в самом начале эксперимента они получали концентрированные сахарные растворы, слегка ароматизированные (например) лавандовым маслом. На расстоянии на 50 м ближе к улью, в веерном порядке были расставлены чашки с нанесенным тем же ароматом, но без пищи. Количество новичков, прибывающих на чашки, соответствовало интенсивности их поиска в разных направлениях. Например, на рисунке 4 показан результат эксперимента, в котором место для кормления находилось в 600 метрах от улья.
Поскольку «веерный эксперимент» доказал, что указание направления было успешным, мы провели «ступенчатый эксперимент» на указание расстояния пчелами. Здесь все ароматизированные пластины были расположены в том же направлении, что и место для кормления, от области улья до расстояния, значительно превышающего расстояние до места для кормления. На рисунке 5 приведен пример эксперимента, в котором место кормления было расположено в 2 километрах от улья. Разумеется, не были оценены прилеты новичков на место для кормления, поскольку здесь был создан дополнительный привлекающий фактор в виде пищи и прибывающих пчел (6).
Подводя итог, этот и предыдущие эксперименты продемонстрировали нам, что информация о направлении и расстоянии до цели была использована с поразительной точностью — и не только в сборе нектара и пыльцы. Те же танцы наблюдаются в рое. Здесь пчелы-разведчицы указывают ожидающим пчелам расположение обнаруженного ими места для поселения. Что наиболее интересно, интенсивность сообщения зависит от качества обнаруженного места для поселения, при этом различные группы пчел-разведчиц конкурируют друг с другом, и поэтому решение, наконец, принимается в пользу лучшего места (10).
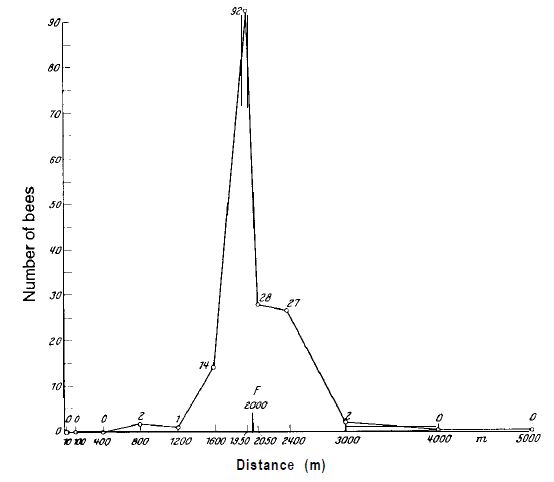
Хотя не вызывает сомнений, что направление и расстояние до цели могут быть определены пчелами по виляющим танцам, группа американских биологов во главе с А. М. Веннером (Adrian M. Wenner) не согласна с тем, что новички пользуются этой информацией. По их мнению, эти пчелы находят цель, используя только обоняние (II). Это мнение несовместимо со многими нашими результатами (6, 12). Оно опровергается приведенным ниже экспериментом.
Пронумерованные пчелы из наблюдательного улья собираются на месте кормления в 230 м от улья. Улей был повернут на бок, чтобы поверхность соты была горизонтальной; небо было закрыто. В этих условиях танцующие пчелы не могли ориентироваться ни на гравитацию, ни на небо, и в замешательстве исполняли танец во всех направлениях. Тарелки с тем же ароматом, что и в месте кормления, располагались на разных расстояниях в направлении места для кормления, а также в трех других направлениях. Их посещали во всех направлениях и в большом количестве новички фуражиры (рис.6), без каких-либо предпочтений к направлению места кормления. После этого наблюдательный улей был повернут на 90° в нормальное положение, что позволяло танцующим пчелам указывать направление к цели на вертикальной поверхности сот. Через несколько минут поток «свежеинформированных» пчел вылетел в сторону места для кормления; ароматизированные тарелки в этом направлении становились все чаще, и за короткое время новички перестали появляться на ароматизированных тарелках в трех других направлениях (рис.7). Не было никаких изменений в источниках запаха в открытом поле или в других внешних условиях. Изменение в поведении новичков-фуражиров могло быть связано только с направленными танцами.
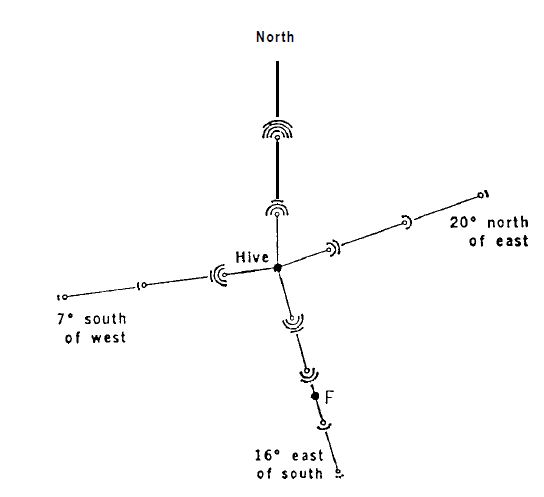
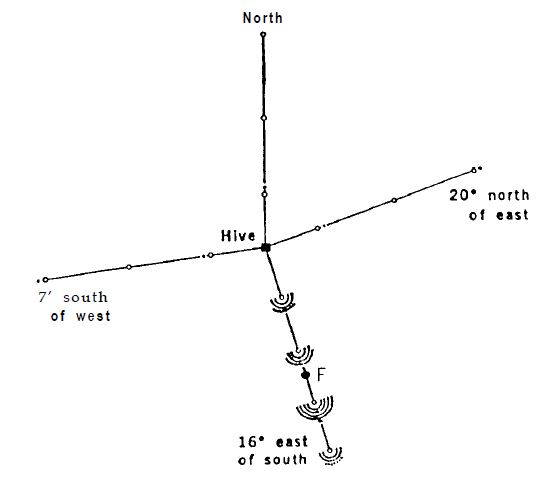
Вполне возможно, что некоторые люди не поверят в это. Лично я тоже вначале усомнился в полученном результате и захотел выяснить, не проявили ли обученные пчелы из моего наблюдательного улья особый поход. Я открыл обычный улей, поднял одну из соторамок и наблюдал за ожидаемыми танцами. С любопытством я повернул соторамку таким образом, чтобы поверхность танца стала горизонтальной. Таким образом, была устранена возможность ориентироваться на гравитацию. Однако, без каких-либо признаков дезориентации, пчелы продолжали танцевать и направлением их виляющих пробегов указывали прямо на место кормления, так же, как мы показываем путь рукой. Когда соторамка была повернута на поворотном столе, пчелы продолжили приспосабливаться к своему новому положению, подобно стрелке компаса (13).
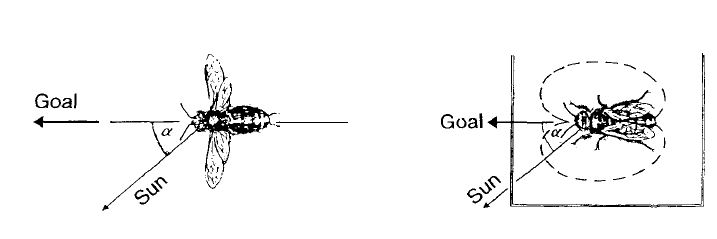
Такое поведение можно изучать на досуге в горизонтальном смотровом улье. В общем, это очень просто, если вспомнить, что направление движения с вилянием брюшком связано с положением солнца. Во время движения по соторамке с вилянием брюшком пчела должна находиться под тем же углом к солнцу, что и при полете в место кормления (рис.8). После того как пчелы-рекруты устанавливают свою траекторию полета под тем же углом к солнцу, они вылетают в направлении цели.
В этом типе дискреционных указаний нет ничего необычного. Прибывающие пчелы-сборщицы нередко начинают танцевать в направлении солнца горизонтальной прилетной доске улья, если их встречают неработающие соплеменницы. Передача информации горизонтальными танцами более понятна, чем при переносе угла на вертикальную поверхность соторамки. Кроме того, у нас, по-видимому, имеется исходный, филогенетически более старый тип указания направления. В Индии все еще существует несколько линий вида Apis. Мой ученик и коллега Мартин Линдауэр (Martin Lindauer) отправился туда, чтобы использовать их для «сравнительного изучения языка». Маленькая пчела, Apis florea, находится на более примитивном уровне, чем наша пчела и другие индийские линии пчел. Колония строит единую сотовую конструкцию на открытой ветке; конструкция имеет расширенный по горизонтали верхний край, который служит исключительно в качестве поверхности для танцев. Когда эти пчелы вытесняются на вертикальную поверхность сотовой конструкции сбоку, они не могут отобразить угол солнца в танце и их танцы с вилянием брюшком становятся дезориентированными (14).
Вернемся к нашим пчелам и понаблюдаем за их танцами на горизонтальном улье. Без сомнения, положение солнца является решающим для направления их танцев. Солнце можно заменить лампой в темной палатке. Изменяя ее положение, мы можем направлять танец пчел в любом желаемом направлении. Однако здесь заключалась большая загадка. Во избежание чрезмерного нагрева во время большинства экспериментов над наблюдательным ульем была установлена защитная крыша. Танцующие пчелы не могли видеть солнце. Тем не менее, в большинстве случаев их танец был правильным. Ориентация по тепловым лучам, проникающему излучению, а также другие объяснения казались возможными и должны были быть отброшены — пока я не осознал, что ориентация по голубому небо и по солнцу в сущности одинаковы. Когда облака покрывали область неба, доступную для взгляда пчел в эксперименте, ориентация танцев немедленно терялась. Должно быть, пчелы могли считывать положение солнца с участка голубого неба. Направление вибрации поляризованного синего света отличается по всему небосводу в зависимости от положения солнца. Таким образом, для того, кто способен воспринимать направление вибрации, даже клочок голубого неба может раскрывать положение солнца по его поляризационной схеме. Наделены ли пчелы этой способностью?
Ответ на этот вопрос был получен в ходе следующего эксперимента. Наблюдательный улей был установлен горизонтально в темной палатке, из которой танцующим пчелам открывался только боковой вид небольшого участка голубого неба. Они правильно танцевали на запад, где на расстоянии 200 метров находилось их место кормления. Когда круглая вращающаяся поляризационная фольга была установлена над соторамкой таким образом, чтобы направление вибрации поляризованного света из того участка неба не менялось, они продолжали свой танец в правильном направлении. Однако при повороте фольги вправо или влево, направление танца пчел менялось вправо или влево на соответствующие угловые значения.
Таким образом, пчелы способны воспринимать поляризованный свет. Небо, которое для наших глаз имеет однородный голубой цвет, для них имеет четкую структуру (13, 15). Они широко используют это и руководствуются в своей ориентации не только положением солнца, но и получающимися в результате поляризационными структурами голубого неба. Они также продолжают распознавать положение солнца после заката или его ухода за гору. И снова пчелы демонстрируют свои чудесные способности. Но теперь ясно, что муравьи и другие насекомые, раки, пауки и даже осьминоги воспринимают поляризованный свет и используют его для ориентации, и что среди всех животных человек вместе с множеством других позвоночных оказался обделенным такими талантами. Однако в одном отношении пчелы остаются уникальными: лишь они используют поляризованный свет не только для собственной ориентации, но и сообщают своим колониям направление к отдаленной цели (6).
Таким образом, язык пчелы, о котором изначально нам стало известно благодаря физиологии чувственного восприятия, теперь вернул нас к этому. Это также подняло общие вопросы ориентации во времени и пространстве. Когда пчелы используют солнце в качестве компаса во время своих полетов, а также для информирования своих соплеменниц, возникает одна сложность: с каждым последующим часом дня положение солнца меняется, и можно предположить, что оно может служить географическим маркером только короткое время.
Я долго размышлял об эксперименте, проведение которого год за годом откладывалось, потому что я не ожидал от него многого. Однако ранним осенним утром в 1949 году мы запечатали вход нашего наблюдательного улья, стоящего в Браннвинкле на берегу Вольфгангзее, перевезли его через озеро и поместили его на расстоянии 5 км в совершенно другом месте, не известном пчелам (15). Пронумерованные пчелы из этой колонии в предыдущие дни посетили место для кормления в 200 м к западу (рис.9).
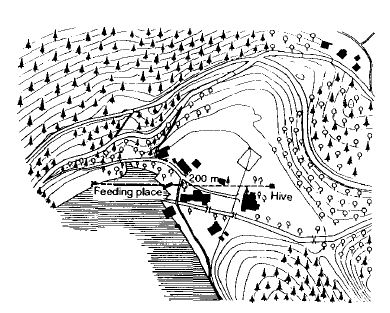
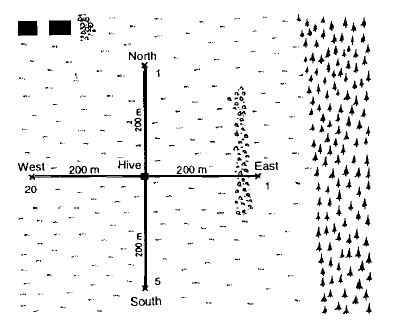
Со знакомого озера и крутых лесистых холмов они теперь оказались на равнинных лугах; ни один из известных ориентиров не были видны. Четыре кормовых чаши с тем же запахом, что и в бывшем месте для кормления, были расставлены на расстоянии 200 м к западу, востоку, северу и югу от улья, после чего вход в улей был открыт. Из 29 отмеченных пчел, которые в предыдущий день в Брунвинквилле посетили место для кормления на западе, 27 обнаружили чаши в течение 3 часов: 5 на юге, по 1 на востоке и севере и, наконец, 20 на западе (рис.10). Все эти пчелы были пойманы после прилета и, таким образом, не могли отправить других, направляя их танцем в улье. Прибывшие пчелы могли ориентироваться только на солнце. Однако солнце находилось на юго-востоке от улья, в то время как в предыдущий день во время последних полетов оно было близко к западному горизонту. Пчелы обладают превосходными внутренними часами. В более ранних экспериментах, когда они кормились в определенные часы, они научились быстро прибывать за стол в это время — даже если тарелка не была установлена. Вышеупомянутый эксперимент, повторенный со множеством отличий в условиях (6, 15, 16), позволил нам утверждать, что пчелам известно ежедневное движение солнца и они могут, вычислив время дня, использовать эту звезду в качестве истинного компаса. Такое же открытие было одновременно и независимо сделано Густавом Крамером (Gustav Kramer) в отношении птиц (6).
В течение последних нескольких лет старый и настойчивый вопрос открыл новую область работы для исследователей пчел. Обсуждая указание направления, изначально я кое-что скрыл от вас. Танцующие пчелы не всегда правильно указывали на источники пищи. В определенные часы они заметно отклонялись влево или вправо. Однако не было никаких неточностей или случайных отклонений; ошибки были согласованными и, когда они регистрировались в тех же условиях, раз за разом давали одинаковые ежедневные кривые. Таким образом, они могли быть правильными, например, для другого положения сот в пространстве. Ошибки возникли только с изменением угла танца; в горизонтальных танцах не наблюдалось «неправильного указания направления». Многолетние наблюдения, проведенные совместно с моим коллегой Линдауэром, наконец, привели нас к выводу, который казался приемлемым (6). Однако он был опровергнут Линдауэром, который упорствовал в своих экспериментах вместе со своим учеником Х. Мартином (H. Martin). Они признали магнитное поле Земли причиной неправильного указания направления. При искусственном экранировании магнитного поля нашей планеты ошибка исчезает; а при искусственном изменении магнитных линий, соответственно менялось неправильное указание направления (17). Идея того, что магнитное поле может играть роль в загадочных способностях животных к ориентации животных, долго отвергалась. В последние годы она подтверждена новыми наблюдениями, особенно птиц и насекомых (18). До сих пор ничто не указывает на возможность того, что пчелы в своих целенаправленных полетах по пересеченной местности используют магнитное поле Земли. Неожиданно, однако, оно оказался столь же биологически значимым, но по другой причине. Когда рой пчел строит свои соты в улье, предоставленном им пчеловодом, их положение в пространстве предписывается маленькими подвесными деревянными рамками. В естественной среде обитания пчелы, например, в дупле дерева, нет никаких деревянных рамок. Тем не менее тысячи пчел работают вместе и в течение одной ночи создают упорядоченную структуру параллельных сот; каждое животное самостоятельно работает здесь и там, не получая инструкций от руководства. Они ориентируются на магнитное поле Земли и едины в определении положения соты, которое они знали из родительской колонии (20).
Хотя решение этих проблем идет полным ходом, едва ли нас ждет множество сюрпризов. Под этим я не подразумеваю, что такие проблемы как восприятие поляризованного света окончательно разрешены. Наоборот: ответ на вопрос, как правило, ставит новые задачи, и было бы самонадеянно предполагать, что окончательная цель будет когда-нибудь достигнута.
В ходе этой лекции не было возможности выйти за пределы краткой иллюстрации, и продемонстрировать ряд важных шагов в развитии наших знаний. Для их подтверждения и расширения требуется больше времени и работы, чем может себе представить сторонний наблюдатель. Усилий одного человека для этого недостаточно. Я нашел помощников и выражаю им свою благодарность. Когда удается найти способных учеников, многие из которых станут постоянными коллегами и друзьями — это один прекраснейших плодов научной работы.
Ссылки и примечания:
1. K. von Frisch, Zool. Jahrb. Abt. Allg. Zool. Physiol. Tiere 35, 1 (1914-1915).
2. ___________,ibid. 40, 1 (1923).
3.___________, Experientia 2 No. 10 (1946).
4. The threshhold of transition from the round dance to the tail-wagging dance varies with each race of honeybees; according to R. Boch [Z. Vergl. Physiol. 40, 289 (1957)], it is about 50 m for Apis mellifica carnica, about 30m for A. mellifica mellifica and A. mellifica intermissa, about 20 m for A. mellifica caucasia and A. mellifica ligustica, and 7 m for A. mellifica fasciala. The fact that the strain we used mostly in our experiments, the Carniolan bee, has the largest round dance circumference was of benefit in our experiments. (Порог перехода от кругового танца к виляющему отличавется у разных видов пчел; в соответствии с R.Boch этот порог составляет 50 м для Apis mellifica carnica, около 20 м для A. mellifica caucasia и A. mellifica ligustica, и 7 м для A. mellifica fasciala. То, что линия, которую мы использовали для экспериментов, принадлежит виду Краинская пчела (Apis mellifera carnica) и имеет самый высокую окружность кругового танца, было удачей нашего исследования)
5. K. von Frisch, Österreich. Zool. Z. 1, I (1946).
6. _________Tanzsprache und Orientienung der Bienen (Springer-Verlag. Berlin, translation: The Dance Language Orientation of Bees (Belknap, Cambridge Mass., 1967)]. Further references are found in this book.
7. H. Esch, Z. Vergl. Physiol. 45, 1 (1961); A. M. Wenner, Anim. Behav. 10, 79
8. K. von Frisch and R. Jander, Z Vergl. Physiol. 40, 239 (1957).
9. I use the word “us,” since the open-field experiments had assumed such proportions that
they could no longer be carried out without trained assistants.
10. M. I.indauer, Z Vergl. Physiol. 37, 263 (1955).
11, A. M. Wenner, The Bee Language Controversy: An Experience in Science (Educational Programs Improvement Corp., Boulder, Colo., 1971).
12. K. von Frisch, Anim. Behav. 21, 628 (1973).
13. ___________,Naturwissenschaften 35, 12 (1948): ibid., p. 38.
14. M. Lindauer, Z. Vergl. Physiol, 38, 521 (1956).
15. K. von Frisch, Experientia 6, 210 (1950).
16. M. Renner, Z. Vergl. Physiol. 40, 85 (1957); ibid. 42, 449 (1959).
17. M. Lindauer and H. Martin, ibid. 60, 219 (1968); M. I.indauer, Rhein. Westjäl. Akad. Wiss. Rep. No. 218 (1971).
18. H. Martin and M. Lindauer, Fortschr. Zool. 21, Nos. 2 and 3 (1973).